ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 05.12.2023
Просмотров: 3450
Скачиваний: 2
СОДЕРЖАНИЕ
Наиболее широко в ЦНС распространены медиаторы - амины:
Другие производные аминокислот - ГАМК, глицин, глютамин и др.
Название рецептора определено медиатором, с которым он взаимодействует:
Взаимодействие гормонов и парагормонов с клетками-мишенями
Понятие высших психических функций (Выготский)
Физиология газообмена в легких
Гуморальная, рефлекторная, нервная регуляция деятельности сердца
1.Общие свойства возбудимых тканей. Процесс возбуждения. Особенности местного и распространяющегося
Механизм формирования ПС связан с:
4. Современные представления о процессе возбуждения. Потенциал действия, его фазы.
8. Механизмы проведения возбуждения по нервным волокнам. Факторы, влияющие на скорость проведения
Механизмы проведения возбуждения по безмиелиновых нервным волокнам такой.
Закономерности проведения возбуждения через нервно-мышечный синапс:
В зависимости от частоты стимуляции выделяют следующие виды мышечного сокращения:
Тетанические сокращения отличается от одиночного следующими параметрами:
12. Функциональная характеристика гладких мышц.
13. Сила и работа мышц. Утомление и его особенности в целостном организме.
14. Нейрон как структурная и функциональная единица ЦНС. Его свойства и функции.
17-18. Возбуждение в ЦНС. Механизмы и закономерности передачи возбуждения в центральных
Особенности передачи возбуждения через центральные аксо-соматические химические синапсы.
Постсинаптическое гиперполяризацийне торможения.
Пресинаптическое деполяризации торможения.
Особенности передачи возбуждения в ЦНС:
Рефлекторная дуга имеет следующие звенья:
24. Рефлекс как элементарный акт нервной регуляции. Строение рефлекторной дуги
По расположению рецепторы подразделяют на:
По виду адекватного раздражителя, воспринимают рецепторы, их подразделяют на:
Физиологические механизмы кодирования информации в рецепторах.
26. Механизм кодирования информации в рецепторах. Адаптация рецепторов.
27. Общие принципы координационной деятельности ЦНС.
28. Суммация возбуждения, торможение нейронами ЦНС. Виды суммации и их значение
В зависимости от локализации рецепторного звена и эффекторного органа рефлексы делят на висцеро-
34. Сегментарные и надсегментарные центры вегетативной нервной системы
35. Гуморальная регуляция, её отличие от нервной. Факторы гуморальной регуляции.
Факторы гуморальной регуляции:
Механизм действия на клетки жирорастворимых гормонов:
Механизм действия жирорастворимых гормонов определяет следующие их особенности:
При воздействии на клетки-мишени водорастворимых гормонов образуются внутриклеточные посредники:
Классификация условных и безусловных рефлексов
- постоянство внутренней среды организма;
Современные представления о путях замыкания временных связей:
Эмоции выполнѐят две функции : сигнальную и регуляторную.
Эмоции делят на низшие и высшие.
Структурное обеспечение эмоций. Эмоциогенные структуры мозга.
Две сигнальные системы действительности
Типы высшей нервной деятельности
Общая характеристика восприятия
Безазотистые органические компоненты крови
Основные физико-химические константы крови:
Противосвертывающая система крови.
В норме гемоглобин содержится в виде нескольких соединений:
Методы исследования вентиляции легких:
Кислородная емкость крови, анализ кривой диссоциации:
Анализ кривой диссоциации НbО2:
^ Рефлекторная регуляция дыхания
второго порядка. Центр второго порядка может вырабатывать 40 - 60 импульсов в минуту.
^ Внутрисердечные механизмы регуляции.
Капиллярный кровоток и его особенности. Микроциркуляция и ее роль в механизме обмена жидкости и
Рефлекторная регуляция сердечно-сосудистой системы в зависимости от изменения положения тела в
Механизмы клубочковой фильтрации. Фильтрационное давление и факторы его определяющего. Состав
Механизм поддержания почками постоянства внутренней среды организма : рН, осмотического давления,
97. Функциональная система питания и пищеварения, ее основные звенья. Сенсорное насыщение. Функции
Пищеварение в полости рта. Состав и физиологическая роль слюны. Слюноотделение, его регуляция
101. Физиологическая роль печени, участие желчи в пищеварении. Факторы стимулирующие секрецию желчи,
106. Физиология щитовидной и околощитовидной желез
107. Физиология надпочечников. Роль гормонов коры и мозгового вещества в регуляции функции организма
Физиологическая характеристика обонятельной сенсорной системы. Механизмы восприятия запахов
Функциональная система –динамическая, избирательно объединенная центрально-периферическая организация, деятельность которой направлена на достижение полезного для организма приспособительного результата.
Свойства, особенности функциональных систем:
1) динамична, способна к постоянной перестройке, к избирательному вовлечению мозговых структур для меняющихся поведенческих реакций;
2) структуры функциональных систем могут быть расположены как на большом, так и на близком расстоянии;
3) вовлечение структур в работу зависит от результата, которого надо достичь;
4) при нарушении функции в одной части системы, происходит срочное перераспределение активности во всей системе;
5) Ф.С. могут устранять, не принимать во внимание те афферентные воздействия, которые не имеют значения для организации конечной приспособительной реакции – принцип сужения афферентации, когда сохраняется только ведущая афферентация.
6) Если конечная цель не достигается, возможно включение дополнительной афферентации.
Функциональная система включает следующие элементы:
· Управляющее устройство – нервный центр;
· Выходные каналы нервного центра (нервы и гормоны);
· Исполнительные органы – эффекторы;
· Рецепторы результата (сенсорные рецепторы) – воспринимают информацию о параметрах процесса и его отклонение от оптимального уровня;
· Каналы обратной связи – с помощью импульсации от рецепторов или на основе изменения химического состава жидкостей информируют нервный центр об эффективности процесса. При этом корректируется деятельность всей функциональной системы.
· Пр: при увеличении давления раздражаются барорецепторы рефлексогенных сосудистых зон, увеличивается поток импульсов в центр кровообращения, изменяется эфферентная импульсация, что ведет к ослабления деятельности сердца и расширению сосудов. Если этого недостаточно включаются дополнительные механизмы – возрастает переход жидкости из кровеносного русла в межклеточную среду и т.д.
Различные константы внутренней среды организма взаимосвязаны – изменение одних констант может привести к изменению других констант. Пр: избыточное потребление воды сопровождается увеличением объема циркулирующей крови.
Все функциональные системы гомеостатического уровня объединяются в функциональную систему гомеостазиса.
Также для регуляции поведенческих реакций, при удовлетворении потребностей организма формируются функциональные системы. Их структуру также разработал Анохин и развивает Судаков К.В.
Структура функциональной системы поведенческого акта:
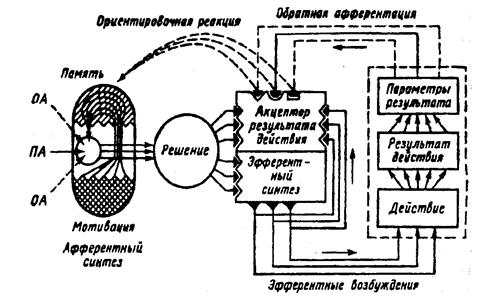
1. Афферентный синтез –процесс взаимодействия возбуждения, возникающего в головном мозге под влиянием обстановочной афферентации (сигналы об обстановке), с возбуждением, вызванным условным сигналом, в результате чего формируется действие.
2. Принятие решений –организм готовится к совершению действия, необходимого именно в данный момент, при этом исключаются остальные неподходящие варианты. Формируется программа действия.
3. Акцептор результата действия –включаются в работу эфферентные структуры, происходит осуществление самого действия. Обратная афферентация сигнализирует о результатах, которые сравниваются с запрограммированными в акцепторе действия. Если цель достигнута система перестает функционировать.
Схема центральной архитектоники поведенческого акта по Анохину П.К
57. Память и ее значение в формировании целостности приспособительных реакций. Особенности механизма кратковременной и долговременной памяти. Физиологические основы обучения.
Огромное значение для индивидуального поведения имеют обучение и память. Выделяют генотипическую или врождённую память ифенотипическую, т.е. приобретённую память. Генотипическая память является основой безусловных рефлексов и инстинктов. Фенотипическая память хранит информацию, поступающую в процессе индивидуальной жизни.
Приобретённая память имеет 2 формы: чувственно-образную и логически-смысловую. Первая формируется в результате действия на анализаторы натуральных раздражителей (запах, вкус, цвет и т.д.), вторая - на основе абстрактных понятий (слова, формулы и т.д.).
Чувственно-образную память делят по характеру раздражителей на зрительную, слуховую, вкусовую и т.д. Обе формы памяти постоянно взаимодействуют, образуя сложные ассоциации (например, название цветка ассоциируется с его внешним видом, запахом). Процесс запоминания происходит в четыре этапа:
1. Сенсорная память. В ней происходит кратковременное удержание сенсорной, т.е., поступившей в органы чувств информации. На этом этапе информация хранится доли секунды. В это время происходит анализ сигналов и большая часть информации переходит в кратковременную память, меньшая - сразу в промежуточную или долговременную.
2. Кратковременная память. Здесь информация находится до несколько минут. Ненужная информация отсюда удаляется, а имеющая значение переходит в промежуточную память.
3. Промежуточная память. В ней информация может храниться от нескольких десятков минут до нескольких лет. Неречевая информация из сенсорной памяти может сразу переходить в промежуточную память (инпринтинг - запечатлевание). Речевая же обязательно поступает через кратковременную память в промежуточную. Причём словесная информация закрепляется в ней лишь после нескольких повторений.
4.Долговременная память. В нее информация переходит из промежуточной, причём этот переход происходит во время быстрого сна.
Первый этап запоминания, т.е. сенсорная память является результатом возникновения нервных импульсов в периферических рецепторах, их распространения по проводящим путям в корковый отдел анализатора и процессов высшего синтеза в коре.
Кратковременная память обусловлена поступлением нервных импульсов в гиппокамп, где выделяется главная и отбрасывается ненужная информация. После этого информация поступает в замкнутые нейронные сети, где происходит циркуляция или реверберация нервных импульсов. Переход информации в промежуточную и долговременную память происходит в коре полушарий на основе более тонких механизмов.
Следы памяти в нейронных цепях коры формируются в результате 2-х процессов:
1. 3а счёт усиления или потенциации нервных импульсов в межнейронных синапсах. Потенциация происходит в результате увеличения количества выделяемого нейромедиатора и числа постсинаптических рецепторов.
2. Благодаря структурным изменениям мембран и органелл нейронов. Эти изменения синаптической передачи и мембран являются следствием предшествующей реверберации.
Данными процессами обеспечивается промежуточная и долговременная память. Кроме того предложены другие теории долговременной памяти.
1. Химическая теория. В её основе лежат опыты с "транспортом памяти" (обучение животных - введение экстракта их мозга необученным животным, опыты со скотофобином). Согласно этой теории информация хранится в специальных белках, синтезируемых нейронами.
2. Теория хранения энграммы в ДНК. Предполагают, что ДНК программирует необходимые изменения структуры и свойств синапсов и таким образом обеспечивает перестройку нейронных цепей в процессе запоминания.
Нарушения памяти;
1. Ретроградная амнезия - утрата способности мозга к извлечению информации, поступившей в мозг до момента экстремального воздействия на него, потеря информации, накопленной до травмы головного мозга или сильного опьянения. Под гипнозом эту информацию можно извлечь.
2. Антеретроградная амнезия - неспособность к запоминанию новой информации. В клинике синдром Корсакова. Память на отдалённые события сохранена, а недавние быстро забываются. Хр. Алкоголизм. Поражение гипнокампа
Каковы же физиологические механизмы, лежащие в основе «кратковременной» и «долговременной» памяти?
Еще в тридцатые и сороковые годы было сделано наблюдение, которое дало основу для того, чтобы высказать гипотезу о природе тех нервных процессов, которые лежат в основе «кратковременной» памяти.
Морфологическими и морфофизиологическими исследованиями американских нейрофизиологов Лоренте-де-Нои Мак Кэллокабыло установлено, что в коре головного мозга существуют аппараты, позволяющие возбуждению длительно циркулировать по замкнутым цепям.Основой служил тот факт, что у аксонов отдельных нейронов существуют веточки, которые возвращаются к телу этого же нейрона и либо непосредственно соприкасаются с ним, либо соприкасаются с отдельными дендритами этого же нейрона; этим самым создается основа для постоянной циркуляции возбуждений в пределах замкнутых круговых цепей, илиреверберирующих кругов возбуждения.Этим простейшим механизмом, однако, дело не ограничивается. Есть все основания думать, что в нервной системе существуют и более сложные аппараты «нейронных сетей», осуществляющих устойчивые реверберационные круги возбуждения. Такими аппаратами являются функциональные комплексы нейронов, соединенные друг с другом «вставочными» нейронами, или нейронами с короткими аксонами, функция которых, по-видимому, состоит в том, чтобы передавать возбуждение от одного нейрона к другому, обеспечивая длительное протекание возбуждения по более сложным сетям, или «реверберационным кругам».
Некоторые исследователи считают, что «реверберационные круги» возбуждения и являются нейрофизиологической основой «кратковременной» памяти. Существенным механизмом сохранения следов оказывается, по этим предположениям, механизм синаптической передачи возбуждения,который и обеспечивает переход возбуждения с одного нейрона на другой и дает возможность осуществлять длительное сохранение возбуждения, протекающего по «реверберациоиным кругам».
Согласно этой теории шок разрушает протекание возбуждения по реверберационным кругам и приводит к исчезновению тех следов, которое сохранялись благодаря этому протеканию возбуждения.
Процесс циркуляции возбуждения по «реверберационным кругам» не является, однако, единственно возможным механизмом сохранения следов. Факты, полученные многими исследователями, заставили предположить, что механизм сохранения следов теми глубокими биохимическими изменениями,которые могут ходить не только в синапсах (места передачи возбуждения от одних нейронов к другим), но и в самих телах нейронов и их отдельных органах (ядрах, метахондриях).
Еще в 1959 г. шведский исследователь Хиден показал, что каждое раздражение нервных клеток приводит к заметному повышению содержания рибонуклеиновой кислоты (РНК), в то время как длительное отсутствие раздражений уменьшает содержание РНК. Дальнейшие наблюдения Хидена и его сотрудников заставили высказать предположение, что изменения РНК носят специфический характер и могут предположительно рассматриваться как биохимический механизм сохранения следов памяти. Основой для предположения является тот факт, что изменения РНК, вызванные определенными воздействиями, могут быть очень специфичны, и что различные воздействия могут вызывать разные модификации РНК.
Было высказано предположение, что число возможных изменений молекул РНК под влиянием различных воздействий измеряется огромным числом – 1015-1020, и, таким образом, РНК оказывается в состоянии сохранять огромное число различных кодов. Как предполагали эти исследователи, повторное появление этого раздражителя приводит к тому, что специфически измененная РНК начинает «резонировать» именно этому раздражению, а способность специфического резонирования именно данному раздражению и является основой того, что нервная клетка, сохраняющая след полученного воздействия, начинает «узнавать» это воздействие, отличая его от всякого другого.
Такое специфическое изменение РНК под влиянием различных воздействий и дало основание исследователям для предположения, что оно является биохимической основой памяти.
Предположение об участии РНК в сохранении следов памяти было подтверждено рядом наблюдений. К ним относятся наблюдения, проведенные известным американским физиологомМореллем,которой показал, что повышение содержания РНК, вызываемое повторным раздражением определенного участка мозга, проявляется не только в этом очаге, но и в симметричном ему пункте другого полушария. Это означает не только то, что реверберационные круги возбуждения могут охватывать очень большие зоны мозга, распространяясь и на противоположное полушарие, но и то, что в этом симметричном «зеркальном фокусе», который не испытывал никакого прямого влияния раздражителя, возникает повышенное содержание РНК, очевидно, указывающее на создавшуюся в нем готовность к повторным возбуждениям.