ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 31.03.2024
Просмотров: 1075
Скачиваний: 0
СОДЕРЖАНИЕ
Математическое моделирование элементов сложных экологических систем
Лекция 1. Введение в моделирование. Исторический экскурс.
1. Основы моделирования в экологии 1.1. Общие принципы построения моделей в экологии
2.2. Этапы построения математической модели
1.4. Элементы теории подобия, применяемые в моделировании
3.2.2. Выборочный метод в экологометрике.
Зависимость числа интервалов от объема выборки
Статистический ряд по интервалам
Лекция 4. Статистические оценки параметров распределения случайных величин по выборкам
4.4. Статистические оценки гипотез об экологических моделях
Выборка из генеральной совокупности
Статистическая таблица эксперимента
Пример преобразования членов уравнения регрессии
Вычисление данных для линеаризации уравнения регрессии
Нормальные уравнения мнк для некоторых функций
Статистическое оценивание уравнения регрессии и парной корреляции.
Обработка результатов наблюдений
Рекомендации по выбору вида функции
3.4. Динамические статистические модели
Данные по объему сброса качественных сточных вод
Данные по объему сброса сточных вод за 5-летие
Пример расчета 5-летних средних
Расчетные значения для определения уравнения динамики
Ряд динамики для определения сезонных колебаний
Эксперименталъный материал исследования
8.1. Анализ влияния отдельных факторов в экологической модели.
Лекция 9. Методы оптимизации. Метод Лагранжа
Лекция 10. Метод линейного программирования.
Лекция 11. Функциональные модели.
Лекция 12. Модели процессов содержащие обыкновенные дифференциальные уравнения.
Численные ошибки использованных для вычисления данных
Лекция 13. Статистические модели динамики.
Лекция 16. Информационные технологии в экологии. Экологические информационные системы.
1 6.1. Экологические информационные системы
1. Какова область значения для числовых характеристик?
Лекция 17. Использование информационных технологий для решения задач экологии.
Критические значения коэффициента корреляции rk;α
2.1. Подобие физических явлений и его признаки
2.4. Применение методов подобия в математическом
11.3. Численные методы решения дифференциальных уравнений
11.3.2. Процесс численного решения
11.3.4. Модифицированный метод Эйлера
11.3.6. Метод Рунге – Кутта для систем дифференциальных уравнений
11.3.7. Общая характеристика одношаговых методов
11.3.9. Методы прогноза и коррекции
11.3.10. Краткая характеристика методов прогноза и коррекции.
11.3.11. Выбор шага и погрешность решения.
11.4. Имитационное моделирование систем
11.4.1. Принципы имитационного моделирования
11.4.3. Динамическая модель исследуемого объекта
11.4.4. Построение имитационных моделей динамических систем
11.4.5. Преобразование передаточных функций звеньев в дифференциальные уравнения в форме Коши
11.4.6. Синтез имитационной модели на основе структурной схемы
11.5. Теоретические основы построения математических моделей систем
11.5.1. Компонентные и топологические уравнения
11.5.2. Компонентные и топологические уравнения механической системы
11.5.3. Компонентные и топологические уравнения электрической системы
11.5.4. Компонентные и топологические уравнения гидравлической системы
11.5.5. Компонентные и топологические уравнения тепловой системы
11.6.1. Сущность метода электроаналогий.
11.6.2. Электромеханические аналогии
11.6.3. Построение имитационных моделей методом электроаналогий
11.6.4. Плоское прямолинейное движение звеньев
11.6.5. Электрогидравлические аналогии
i*
Для перехода к новой симплекс-таблице необходимо поменять места- ми х1 и х4, а все элементы разрешающей строки умножаем на элемент а*41 = 1. Все элементы разрешающего столбца умножаем на -а*41=-1, кроме самого разрешающего элемента, что приведено в табл. 5.3.
Таблица 5.3
j*
|
|
х1 |
х2 |
х3 |
ьj |
|
х4 |
1 |
1/2 |
1/2 |
5/2 |
|
х5 |
-1 |
3 |
-1/2 |
4 |
|
х6 |
1 |
2 |
1/2 |
8 |
|
pj |
-3 |
1 |
3 |
F* |
i*
Остальные
элементы
таблицы
вычисляем
по
формуле,
значение
нового
элемента
![]() равно:
равно:
![]()
Тогда для элемента а52
![]()
Аналогично для рj
![]()
и для bi
![]()
Так же для F
![]() и т.д.
и т.д.
Заполняем табл. 5.3. Так как все коэффициенты рj в табл. 5.3 положительны, решение найдено. Значение F из последней строки табл. 5.3 с противоположным знаком будет решением целевой функции, т.е.
Fmin = — 7.
Базисное решение: х1 = 2,5; х2 = 0; х3 = 0;
x4=0; х5 = 6,5; х6 = 5,5.
Проверка: Fmin= -3х1 + х2 + 3х3 = -3 х 5/2 = -7,5.
назад
Лекция 11. Функциональные модели.
Функциональная зависимость может выражаться в виде алгебраических равенств или дифференциальных уравнений. Рассмотрим наиболее типичные функции, моделирующие те или иные экологические процессы.
Функции роста чаще всего обозначают некоторую аналитическую зависимость между ростом параметра, например массы вещества, и факторами, обусловливающими этот рост. Так, если рас- сматривать прирост сухого вещества растения W в зависимости от прироста субстрата S за период времени dt, то получим уравнение роста
![]()
Далее темп роста (при условии, что система не получает из внешней среды и не теряет никакого материала) можно предста- вить в виде функции
![]() .
.
Если принять допущения, что количество энергии роста пропорционально сухой массе W; механизм роста имеет максимальный темп на протяжении всего времени, пока существует питательная среда; процесс роста необратим и прекращается, как только истощается питательная среда, то
![]()
где μ — удельный (относительный) темп роста.
Параметр μ , зависит от вида сухой массы W соответствую- щей ресурсу питательной среды, и от скорости, с которой работает механизм роста. Интегрируя выражение (5.9), получим:
W= W0е?t, при 0 < t< tf,
W=Wf, при t>tf.
Когда W = Wf, S = 0, то Wf = W0 + S0 и рост внезапно прекращается, тогда
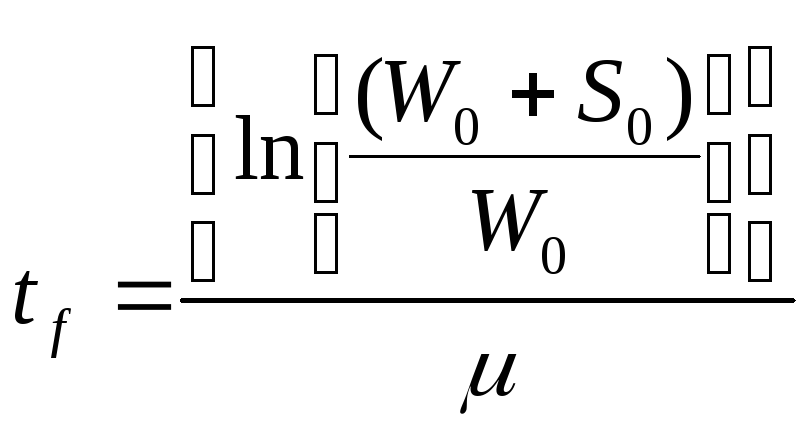
(здесь приводится результат подстановки Wf = W0 + S0 и t = tf ).
Простой экспоненциальный рост, ограниченный реальным ресурсом питательной среды, графически представлен на рис. 11.2.
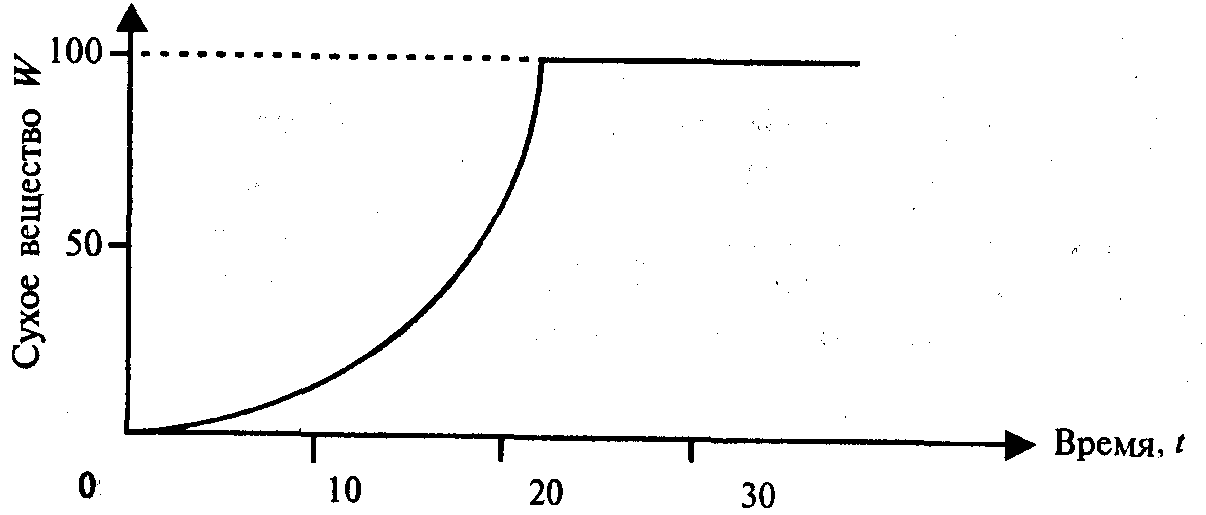
Если предположить, что механизм роста работает со скоростью, пропорциональной ресурсу питательной среды S, то можно записать:
![]() ,
,
где k — некоторая постоянная величина.
Тогда при S= Wf — W, получим:
![]()
Интегрируя
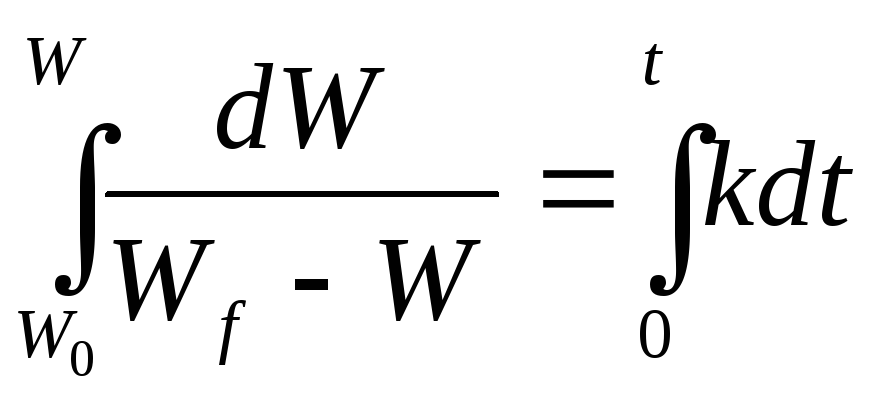
получим
![]() .
(11.10)(5.10)
.
(11.10)(5.10)
При W0 = 0
W = Wf(1 — е-kt).
Зависимость 11.10(5.10) графически представлена на рис. 11.3. Она представляет собой мономолекулярный рост: уравнение описывает ход простой необратимой химической реакции первого порядка.
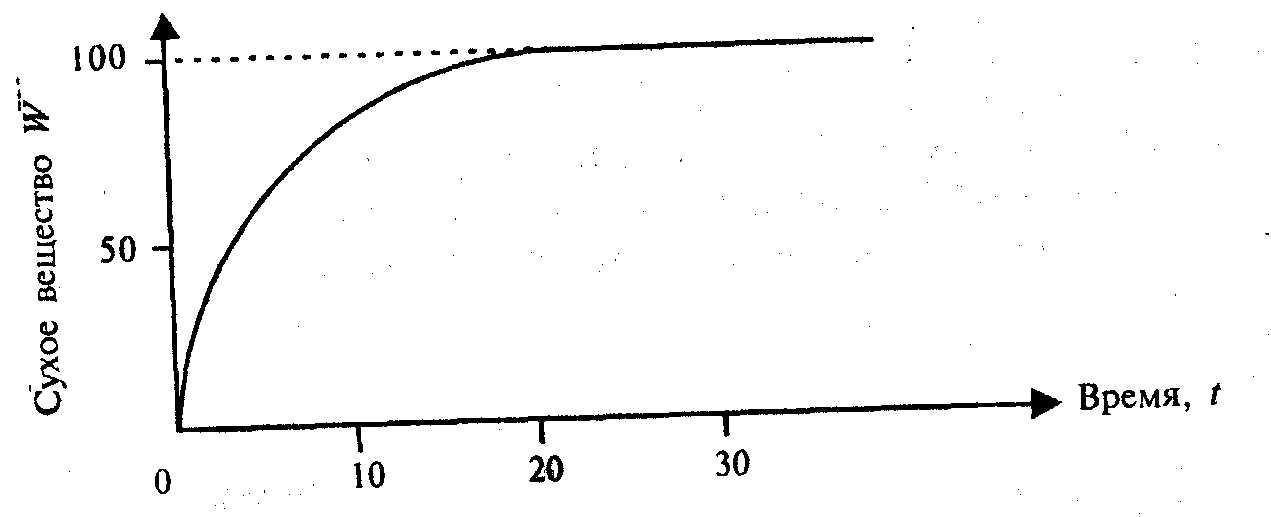
Далее можно показать, что уравнения роста могут быть:
• логистические
![]() '
'
• Гомперца
![]() ,
,
где D — дополнительный параметр, характеризующий уменьшение μ;
• Ф. Ричардса
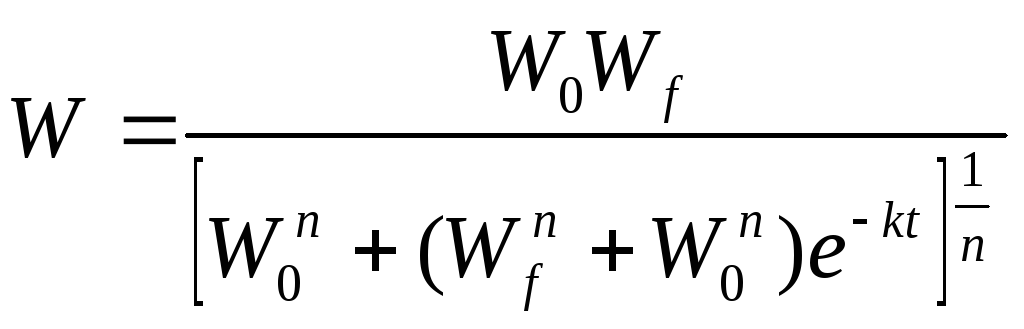
где n — параметр, n ≥ - 1;
• Л. Чантера
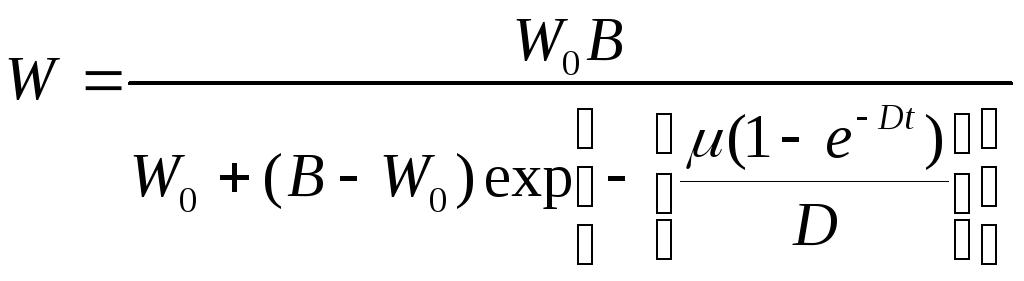
где В и D — постоянные величины.
Д. Хаксли впервые была описана аллометрическая зависимость для описания роста животных и частей их организма, а затем В. Перселлом эта зависимость была использована для описания роста растений.
Предположим, что P и Q — некоторые свойства организма (на- блюдаемые количественные характеристики): например P и Q, мо- гут быть массами различных частей животного или Р может зада- вать сухую массу растения, а Q — площадь поверхности его листь- ев. Поскольку организм растет и развивается, то P и Q будут изменяться с течением времени, т.е.
Р≡ P(t); Q ≡ Q(t).
Считается, что Р и Q аллометрически зависимы, если они удов- летворяют аллометрическому уравнению
р =аQb (11.10)(5.11)
где а и b — постоянные коэффициенты.
P и Q изменяются во времени таким образом, что уравнение (11.10)(5.11) сохраняет справедливость во всем интервале наблюдений.
При
![]() и
и
![]() можнозаписать
можнозаписать
Q= сРd.
Логарифмируя (5.11), получим:
ln P = lnа + blnQ
и, дифференцируя по времени
![]()
Таким образом, пропорциональные темпы роста зависят от постоянного фактора b. Аллометрическое уравнение нашло применение во многих задачах. Так, в форме
W1=![]()
оно описывает отношение сухой массы корней к сухой массе побегов в растущем растении. Многие модели роста могут быть представлены в форме
![]()
где W — масса;
t - время;
Р — множество параметров или постоянных коэффициентов, характеризующих организм;
Еm — параметры или переменные, характеризующие те условия окружающей среды, которые поддаются управлению;
ЕW — показатели погоды.
Во многих случаях в экологии могут быть использованы модели «хищник — жертва». Впервые эти вопросы были затронуты в работах А. Лотки и В. Вольтера.
Пусть для данной естественной среды у обозначает число жертв, а х — число хищников. Согласно Вольтеру, имеет место:
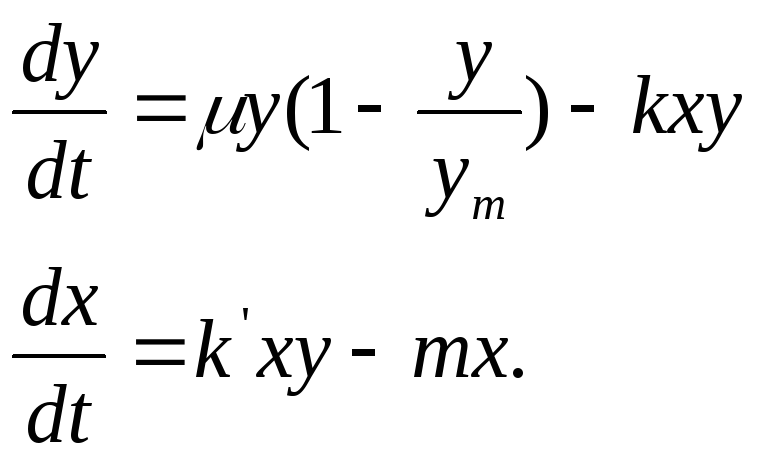 (11.12)(5.12)
(11.12)(5.12)
где μ, k, k’, t — параметры;
уm — число жертв (1/сут.).
Уравнения (5.12) могут дать решения:
1) для автономного роста хищников